莫里遜層有著高多樣性的蜥腳類恐龍。由於資源有限,為了不吃垮生態系,這些巨龍便有了食性上的差異與行為上的適應。本篇介紹巨龍間的資源利用。這類資訊過去很難直接從化石取得,但拜新科技之賜,隱藏在化石裡的秘密逐漸被揭開,特別是牙齒。

化石裡隱藏的龍生
要研究化石動物的食性分化在過去有相當的難度。直接證據主要來自如糞化石、嘔吐物等痕跡化石或化石上的齒痕,但很難確認真正的產出者,只能從尺寸、內含物與當地的化石物種綜合推測;最好的直接證據是骨架內的胃含物,但能如此保存下來的化石少之又少;拜現代科技之賜,科學家已能對實體化石本身做更精細的觀察,或對外觀無法直接觀察到的部位做檢視,藉此蒐集相關的生態資訊,甚至進一步以電腦模擬,做出間接但具有證據力的推論。而取食的第一關嘴巴,無疑隱藏著許多重要線索。

知齒知彼
先說牙齒的形狀和分布。雖然巨大的蜥腳類恐龍乍看都一個樣,但細看的話牠們的頭型和牙齒卻有很明顯的差異。
例如梁龍科的梁龍、迷惑龍等都具有相對較扁平的頭和寬闊的嘴,如鉛筆狀(或稱木樁狀)的長牙只分布在嘴巴的前端,適合將植物切斷或扯斷,但無法咀嚼;圓頂龍一如其名,頭型較圓,牙齒粗短,狀如抹刀或湯匙,分布在整個上下頷,能上下咬合,有限度地處理口中的食物;腕龍的頭型更高,牙齒與圓頂龍類似,雖然沒那麼粗壯,但數目更多。模擬重建上下頷肌肉和咬合時的應力測試,也顯示兩者有明顯差異,梁龍的咬合力遠不如圓頂龍。
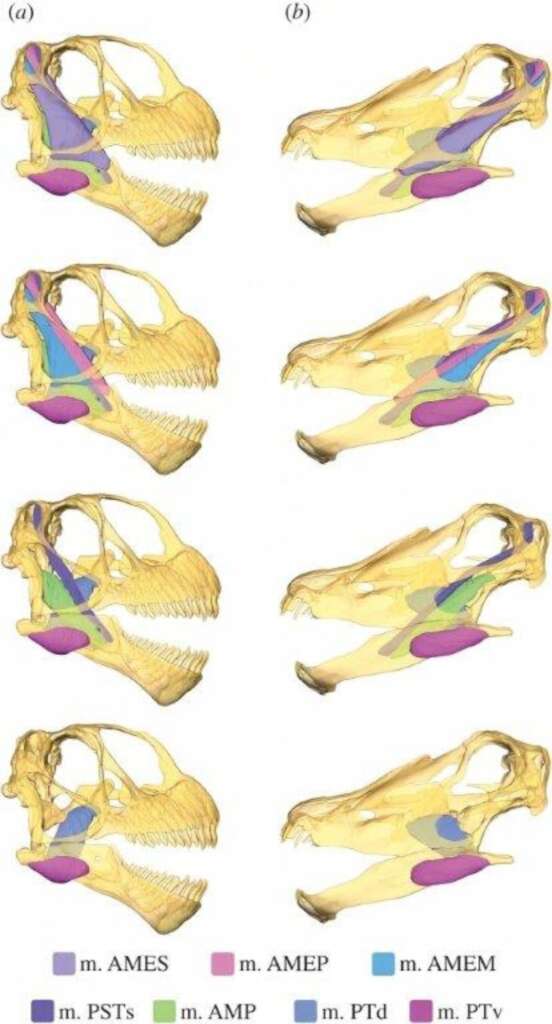
牠們的牙齒齒冠琺瑯質的磨損也出現有趣的型式,梁龍類通常磨損痕跡較細緻,而在圓頂龍與腕龍則粗糙許多。古生物學家對此有兩種不互斥的解釋,一是前者取食較柔軟的植物,後者則吃比較堅韌粗糙的植物;二是前者的取食高度較低,容易吃到地面的沙塵而出現細微磨損,後者則否。近年根據頸椎細部結構和力學模擬的骨骼重建,主張梁龍科成員的脖子雖長,但慣常姿勢是水平前伸或略微抬高,和鞭子般的長尾巴約略呈水平或傾斜直線。圓頂龍與腕龍則平常是脖子傾斜或高舉的姿態,尾巴離地,這也符合前述的推論。

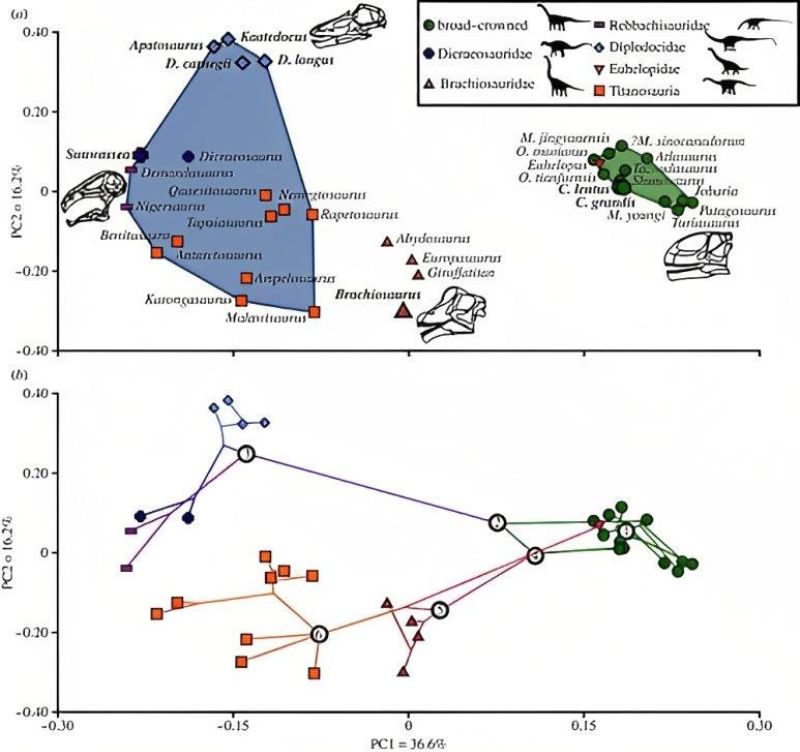
綜合上述,古生物學家推測梁龍科的取食行為比較像是水平吊臂,身體不動的情況下左右橫移脖子取食較低矮的木賊蕨類等較柔軟的植物;圓頂龍則是靠著高脖子取食針葉樹較堅韌的枝葉;身體更高的腕龍甚至能取食樹冠層。如此使得同一地區不同科的恐龍在食物上有所分化,也有偏好的取食環境:前者偏好較開闊的環境,後者則待在樹林內取食。而根據功能性空間分析,莫里遜層的主要蜥腳類恐龍也有明顯的分群。因此在形態構造、磨損、生物力學等證據上,都支持牠們在食性上有所區分,而這些區分或多或少跟牠們的譜系關係有關。
另有玄機
除了上述這些形態上的分化,還有一些不那麼直觀的跡證,必須透過精密儀器或是串聯不同標本才能發現。比如骨頭底下的構造和發育上的改變。
現生有蹄類哺乳類的牙齒有分化,前端齒用來切斷植物,後端齒用來咀嚼,中間還有間隔可容納食物;恆齒具有厚厚的琺瑯質,較耐磨損。反過頭來看,蜥腳類的牙齒既簡單也無分化,主要用來切斷或扯斷植物,不利咀嚼,且琺瑯質相對薄,很快便會磨損。對此,蜥腳類發展出草食獸演化史上很獨特的應對方式:快速換牙。

科學家掃描迷惑龍和圓頂龍等的頭骨,發現每顆牙的齒槽裡都還藏有許多備用牙,最多可達 8顆,最少也有 3 顆,迷惑龍的數量又比圓頂龍多。根據牙本質(dentine)的發育速度推測,前者平均大約每個月就能換牙,而且是整批換掉而非每次單顆替換,後者則約兩個月。現今多數哺乳動物的恆齒長滿後便不再換牙(例外如大象),反觀蜥腳類恐龍,是以低品質的牙齒搭配高速率換牙來因應磨損。
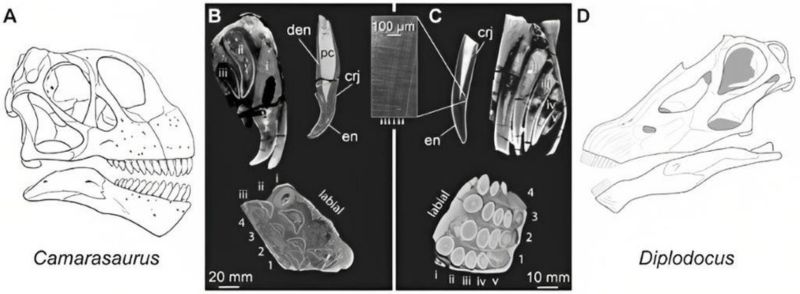
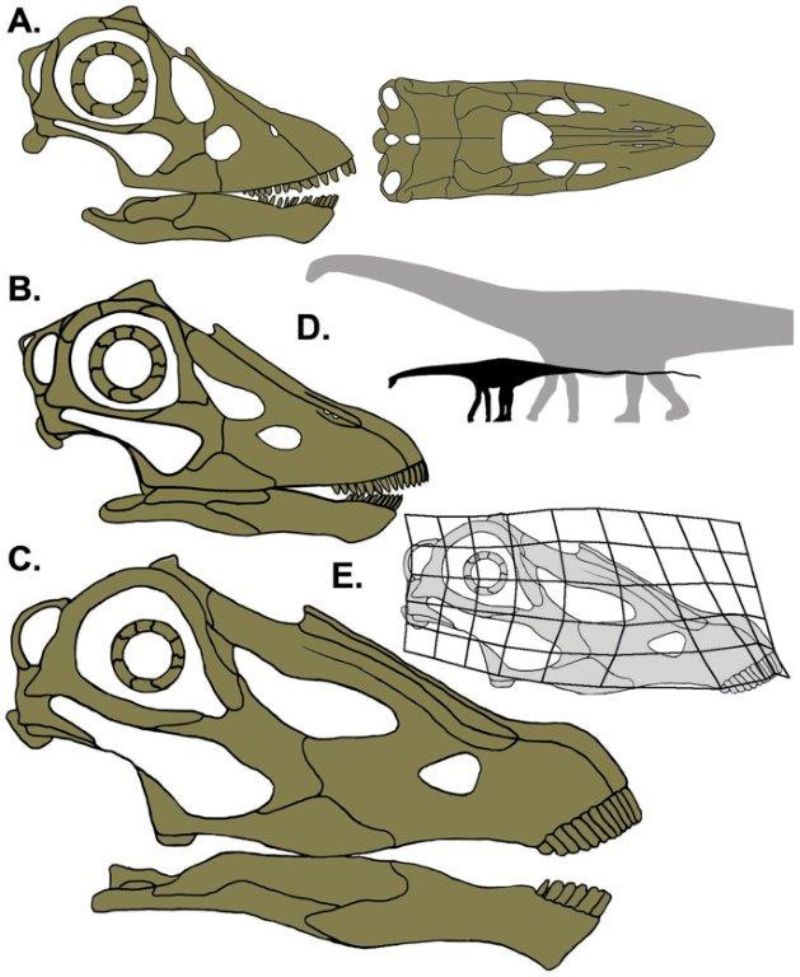
2010 年在蒙大拿州的母親節挖掘場(Mother’s Day Quarry)挖到一個部分完整的小頭骨,而當地出土的清一色是小型的梁龍類骨骸,也許都是幼體。這個頭骨長度只有 24 公分,約成體的一半大小,是目前發現最小的梁龍頭骨。當地另出土過 3 件大小不一的頭骨標本,正好可以用來重建發育過程的變化。
這在 2018 年有了有趣的發現:幼年梁龍的頭型不像成體那麼扁,吻部比較短圓,牙齒分布不那麼趨向前端,且牙齒的形狀和磨損狀態更類似圓頂龍。圓頂龍是保有比較多祖徵的蜥腳類(所謂的「原始」),換句話說,幼年的梁龍也保有較多祖徵,在爾後的發育過程中逐漸出現梁龍特化的衍徵,這與現今演化的概念相符。而這些差異也反映出梁龍幼體與成體之間應該有食性和棲息環境上的轉化。

但無論何者,這些蜥腳類恐龍還是有著眾多的先天限制,只能不經或做有限的咀嚼後便直接把食物吞下肚,再借助胃石幫助消化。加上牠們無法咀嚼,肯定不會反芻。如此使得牠們在單位時間內能攝取的營養和能量相對較低,因此每天需要長時間進食,吃進很多食物。可是一個地方真有那麼多食物可以吃?
逐水而居
文章開頭已講到,莫里遜盆地並非一個資源豐富的環境,季節性濕潤的半乾燥氣候很難維持植物長時間的生長,跟東非的稀樹草原類似。但後者卻餵養著巨量的草食獸,當中的生態機制便是跟著雨季循環的草食獸的大遷徙,讓土地可以漸次休養生息恢復生產力。
莫里遜層可能也有類似的生態機制,境內留有許多恐龍行走足跡的生痕化石,遺跡多達六十餘處。最大的一處有 1900 多個蜥腳類、獸腳類、鳥臀類留下的足印,但以蜥腳類為大宗。有趣的是,蜥腳類的足跡通常具有約略平行的方向性,且大小足印都有,顯示可能為成群往某個區域移動;相對地,獸腳類恐龍的足跡則無此現象,且較零星,顯示是單獨追獵或在棲地四處移動所留下。鳥臀類足跡相對稀少,無法判別是否會跟蜥腳類成群遷移。

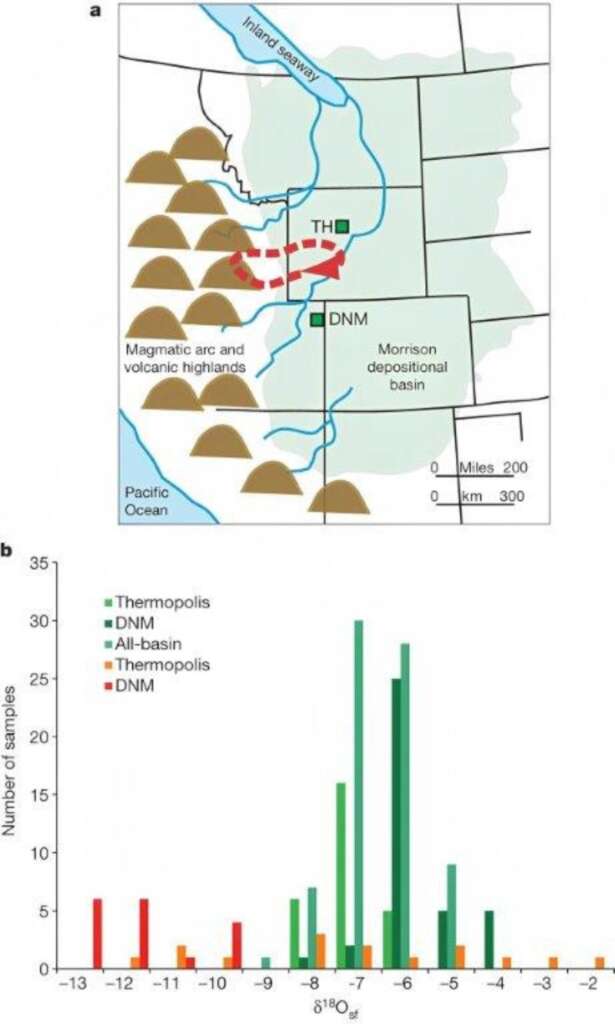
另一個更強的證據是來自牙齒。動物喝水時,會將當地水源中的氧 18 穩定同位素(18O)攝入體內。由於水中的氧也被用於生成碳酸鈣(CaCO3),因此像淡水雙殼貝或螺的殼便會記錄下地表水 18O 的分餾比值(δ18Osf),而每個地區的 δ18Osf 都有所不同。牙齒的琺瑯質同樣是含氧化合物,生成過程中也同樣會記錄當時的 δ18Osf 。若化石發現地的背景 δ18Osf 與琺瑯質的不同,可能就意味著動物的長距離移動。
在這個邏輯推理下, 2011 年果然在圓頂龍的牙齒上發現遷移的證據。被研究的牙齒分別來自懷俄明州瑟莫波里斯(Thermopolis,TH)與猶他州的國立恐龍公園園區(DNM),兩處都位於盆地的低平地區。源自DNM的牙齒,其琺瑯質的 δ18Osf 顯著低於背景值, TH 的變異則很大,但與背景值重疊。這顯示前者在牙齒生成時並不在 DNM ,而可能是在盆地西側的高山區,因為 18O 比較重,水氣中的 18O 多半會在半途被下掉,導致高山水源的 δ18Osf 偏低,莫里遜盆地其他地點的 δ18Osf 則趨近平均值。 DNM 牙齒的主人回到盆地後死亡被埋藏,顯示了長距離的移動。至於 TH 的牙齒主人,可能就地待了相當長的時間,因此琺瑯質的 δ18Osf 並未記錄到遷移的證據。
若一切假設為真,則檢測的結果強烈支持圓頂龍會做長距離的遷移,單向的距離可能達 300 公里或更遠。原作者推論這樣的長距離移動是季節性遷移,以牙齒生成時間來看,可能在山上待約 5-6 個月。但前面提到的 2013 年研究主張圓頂龍的預備牙約在 2 個月便可生成,因此只能推測牠們至少會在山上待兩個月,也至少會在平地待兩個月,但真正的遷移週期還需要更多牙齒樣本才可能回答。


本著作由本館研究人員所提供,博學多文團隊編輯製作,以創用CC 姓名標示–非商業性– 禁止改作 4.0 國際 (CC BY-NC-ND 4.0) 授權條款釋出。若需要使用本篇的文字、圖像等,請洽本館出版室。