
「Legend of Cretaceous丨白垩传○2」末世对决(下):角龙的反击

■宿命:角龙超科篇
| 前 言
得益于大量全新开花植物的繁盛,此时的地球逐渐变得比以往任何时期都更加斑斓绚丽。
直至坎潘阶时,不断涌现的被子植物已成为优势种群并伴随着蜜蜂等昆虫的协同演化彻底改变了地球植被面貌:榕树、悬铃木、木兰花、蔷薇等大型植物展露头脚并一直盛放至今;一些大型的裸子植物如南洋杉、松柏目、苏铁等也依旧生机勃勃的广泛分布于全球各地;“活化石”银杏更是随处可见;自侏罗纪时期就繁茂无比的羊齿类、木贼等植物到了中生代末期也并未见衰落,新旧并存野蛮生长着。








与此同时,曾经的盘古大陆在早白垩纪便分崩离析,这也使得各大板块之上的物种逐渐开始独立演化,进一步促成了各大种群日渐多样化并呈现出明显的“南北差异”:
在北半球的北美和亚洲,兽脚亚目的代表以暴龙科和众多中小型驰龙科、伤齿龙科、偷蛋龙下目为主,优势性的植食性动物则被角龙科、甲龙科、鸭嘴龙科和厚头龙下目等鸟臀目演化支共同占据,且伴随着首批禾本科植物(草)的辐射演化,更加衍化的鸭嘴龙科和甲龙下目得以成为最成功、适应性最强的植食性动物,种群多样化达到极致。
而兽脚亚目在这样的新趋势之下也不断出现杂食性的种群,如镰刀龙超科和似鸟龙下目,不断拓宽种群生态位。






在一端的南半球,冈瓦纳大陆依旧在不断持续分裂中,这里的生态面貌与北半球截然不同:阿贝力龙科成为了当地最优势的大型兽脚亚目掠食者,而鹫龙、半鸟等中小型驰龙科也分布广泛;蜥脚亚目在这里找到了最后的乐土,它们依旧繁盛无比并演化出史上最叹为观止的陆地巨兽如泰坦巨龙、阿根廷龙。






各种开花植物的持续扩散演化也使得不同种类的植食性恐龙发展适应出各色差异化的进食方式,如:角龙类演化出如同鹦鹉一样的喙用于植物果实和根茎进行切割,而后方众多的齿系则可以咀嚼。
这种针对咀嚼能力出现的齿系(tooth batteries)在鸟臀目的其他演化支上都有出现并被鸭嘴龙科发展到极致(类似齿系在部分蜥脚亚目如雷巴齐斯龙科身上也有发现),它们趋同演化出和哺乳纲偶蹄目一样的咀嚼功能,尽管两者并无关联属于典型的非同源平行演化。这在几乎都采取“囫囵吞枣”模式的蜥形纲中,算得上是绝无仅有的特例。


而以角龙科和鸭嘴龙科为代表的鸟臀目恐龙对新类型被子植物的快速适应性演化或许正是它们能够成为后起之秀崛起,并最终得以在白垩纪晚期赶超了大型蜥脚亚目霸主地位的关键因素。
这其中,角龙科作为白垩纪演化辐射最成功、种群多样化最明显的动物,同暴龙科一样都是指标性的明星物种。两者一攻一守,共同上演了一出延烧至末世余辉前的漫长拉锯战。
接下来,就让我们来看看面对咄咄逼人不断进击的暴龙,角龙又是如何步步为营予以反击的。


“地表最强防御者”
同暴龙超科一样,角龙超科(Ceratopsoidea)也是一个比较庞杂的概念,由奥赛内尔.马什(Othniel Marsh)在1888年首次提出并使用,是一个隶属于鸟臀目角龙下目之下的一个超科,其中最明显的外观特征就是具有角和头盾。
它们普遍生存于白垩纪土伦阶-马斯特里赫特阶之间(约9400~6500万年前),是白垩纪-古近纪大灭绝事件前存活时間最晚的几种恐龙种群之一。
●【演化谱系】:蜥形纲-恐龙总目(Dinosauria)-鸟臀目(Ornithischia)-角足亚目(Cerapoda)-角龙下目(Ceratopsia)-新角龙类(Neoceratopsia)-角龙超科(Ceratopsoidea)
【生存年代】:距今9400—6500万年前,白垩纪土伦阶-马斯特里赫特阶;主要分布于北美洲。
被誉为“最强陆地植食性动物”的角龙科在恐龙演化史上究竟处于怎样的位置呢?要回答这个问题就需要了解一下鸟臀目的具体分类。
恐龙总目作为一个异常庞杂的演化支被划分为蜥臀目和鸟臀目两大阵营,两者很早就分道扬镳各自独立演化。相对于蜥臀目旗下囊括了各大肉食性种群的兽脚亚目演化支和后裔鸟类以及巨兽层出不穷的蜥脚亚目,鸟臀目多少显得有些黯淡,但其实它们也不乏一些明星物种。
与蜥臀目两大阵营一样,鸟臀目也下分为装甲亚目和角足亚目两大演化支,它们分别代表了鸟臀目往不同方向进行防御性演化的尝试:



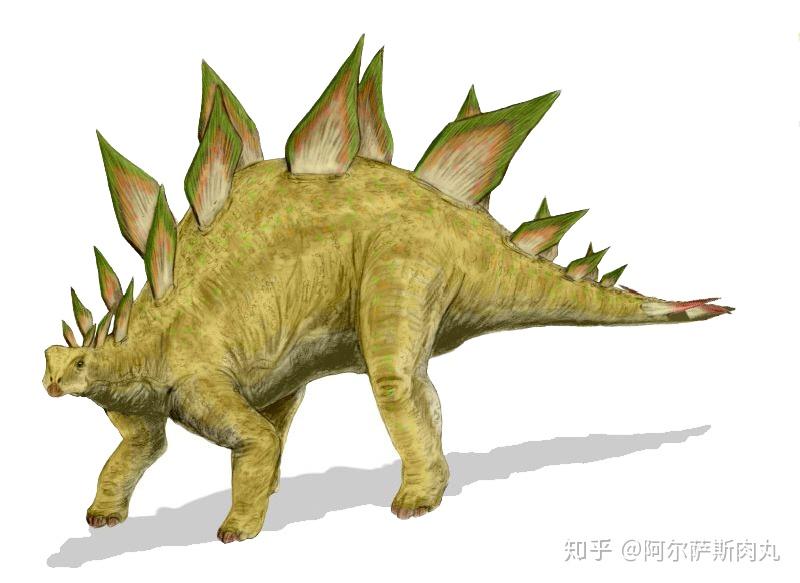
其中一支主要靠强化背部和尾部的装甲、骨板、骨刺等装备用于抵御来犯掠食者的装甲龙类就包括了大众耳熟能详的剑龙下目和甲龙下目;另一支则是不断强化头部头饰、尖角、骨盾为主的角足类。
根据外观和体征结构的差异又可以将它们细分为鸟脚下目和头饰龙类,前者包括了高度衍化极为成功的鸭嘴龙超科;后者则包含了我们今天的主角——角龙下目和在《侏罗纪世界2》中出尽风采的厚头龙下目。
由此看来,暴龙与三角龙之间的恩怨情仇也算得上是蜥臀目-兽脚亚目最强掠食者和鸟臀目-角龙下目最强防御者两大阵营间的“终极对决”。当角龙科的防守之路还在起步之时,最早的暴龙超科成员也已在亚洲大陆出现,两个物种间的攻防战就这么悄然拉开序幕。
“燎原”
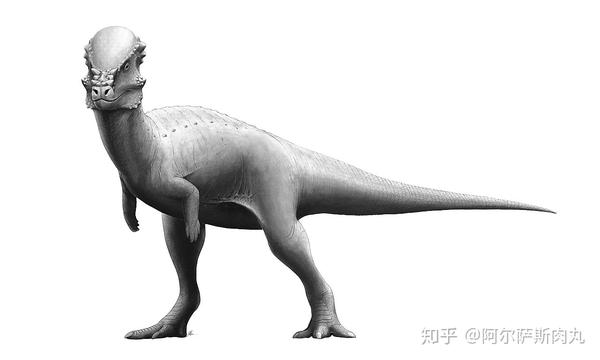
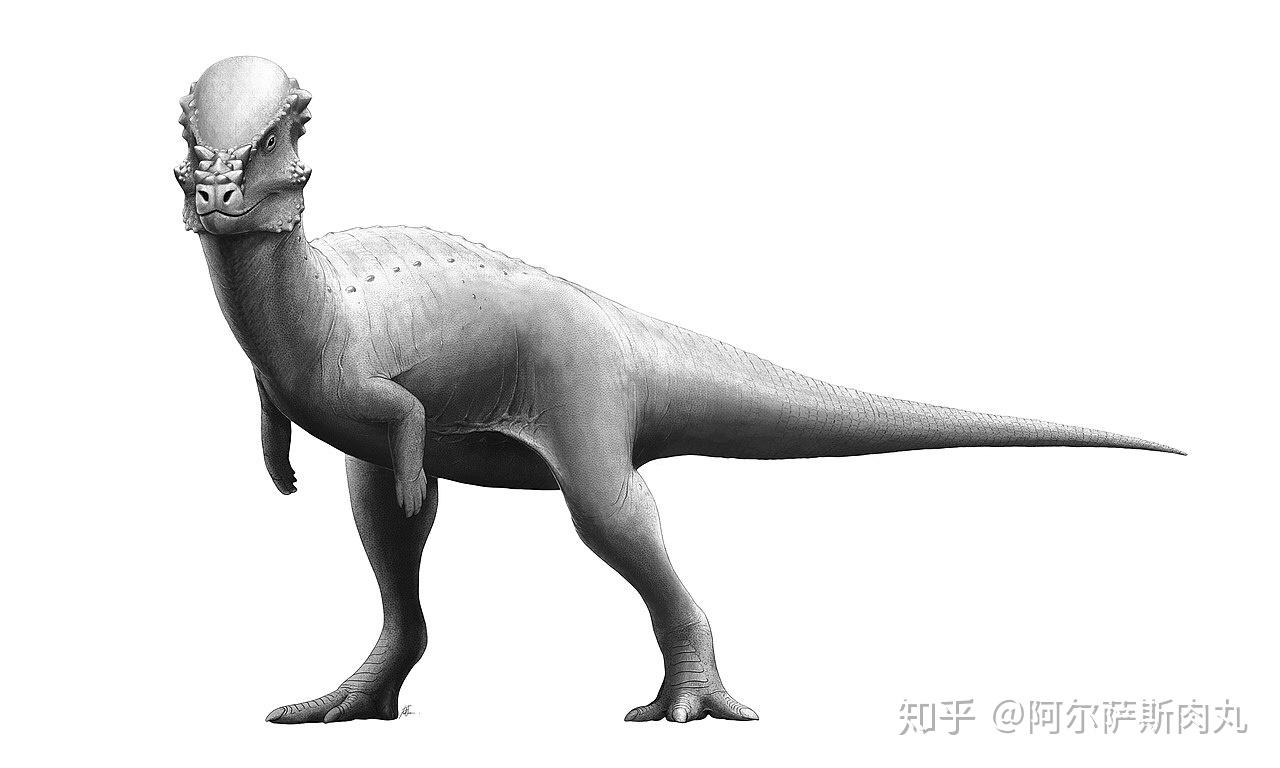
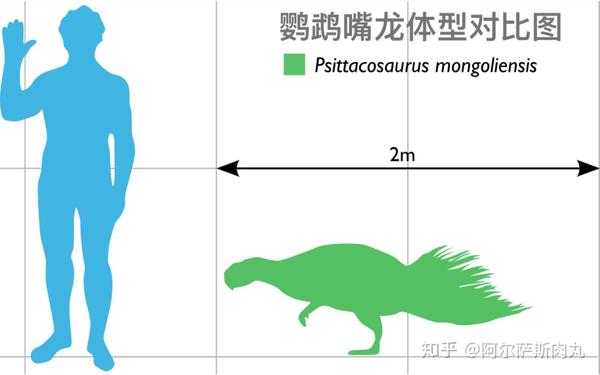
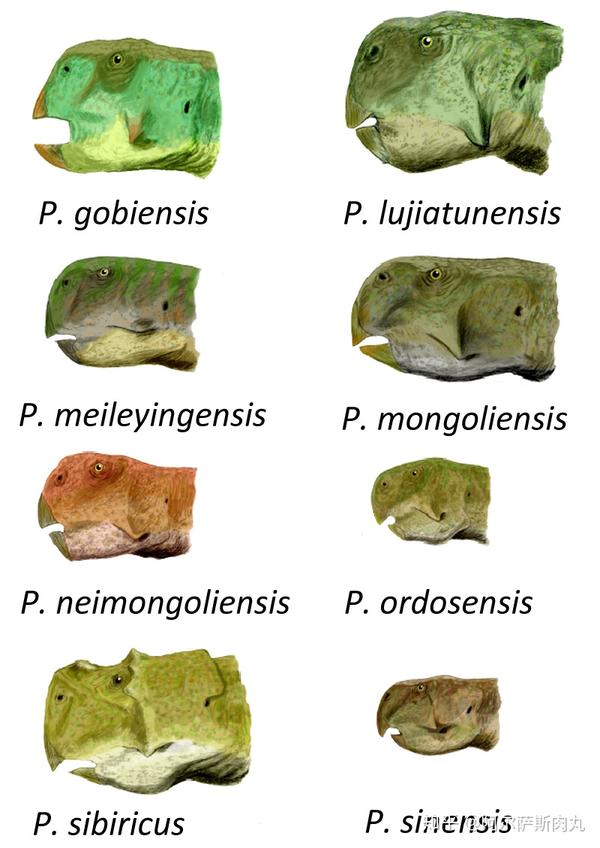
●【鹦鹉嘴龙科 Psittacosauridae】:拉丁学名是“鹦鹉蜥蜴”之意。
【生存年代】:早白垩纪(距今1亿4020万年-9960万年前);蒙古国、中国、俄罗斯。其中包括9-11个有效种。
同暴龙超科早期的成员一样,最原始的角龙类也是一群体型娇小的两足类恐龙,与后期进阶并高度衍化拥有如犀牛、大象一般身躯的后辈相比,处于早期起步阶段的角龙并不出众,而鹦鹉嘴龙便是这一阶段的代表性物种。
不过需要明白的一点是,鹦鹉嘴龙并非是后来一系列大型角龙科的直系祖先,只能算是它们的近亲。

这群大小类似瞪羚、两足步态、带有喙状嘴、尾部长有鬃毛的小型植食性恐龙是非常早期的角龙下目成员,尽管在体型和外观上同晚期的角龙类有着诸多差异但它们身上依旧具备不少后期进步型角龙的共有衍征。
虽然没有同门的三角龙这么高的大众知名度,但鹦鹉嘴龙作为目前为止发掘出的最完整的恐龙之一(目前已发现了400多个从幼体到成体的完整骨骸化石),使得科学家研究其成长速率和角龙科成员的演化路径提供了非常直观和可靠的依据。而它们数量庞大的化石记录,更是成为判定早白垩纪沉积层中的指标。
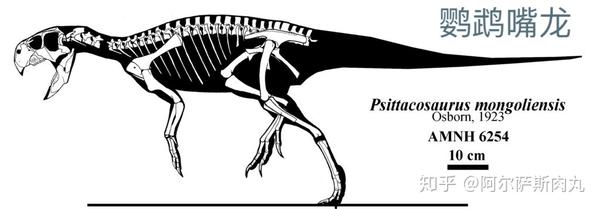

顾名思义,鹦鹉嘴龙的学名来源正是它们那趋同于鹦鹉的角质喙嘴。它们的上下颌分别由喙骨(rostral bone)和前齿骨形成,喙嘴外部被角质包裹形成锐利的切割面便于切碎咬破植物根茎和种子外壳,其功能和现代的鹦鹉喙如出一辙。
观察鹦鹉嘴龙的颅骨可以发现,它们也有许多与更衍化的角龙科相同的适应性衍征:上颌前端的独有喙骨、朝两侧突出的颧骨。不过它们尚未发展出晚期角龙科那样夸张的头盾和角状结构。
与绝大部分早期的鸟臀类一样,鹦鹉嘴龙是典型的两足直立步态。纵观整个角龙类的演化史我们可以清晰地看到这样一个趋势:伴随着体型的不断增加、消化系统的强化和生态位的持续特化,角龙科从早期的两足步态逐渐演化为四足步态。而这种伴随体型的增加从两足转变为四足步态的例子在很多植食性恐龙大型化的演变之路上也算是屡见不鲜了(蜥脚亚目、鸟脚下目同理)。






鹦鹉嘴龙的前肢长度比例仅有后肢的58%根本无法触碰到地面,再根据其前肢活动范围可以推断出它们不具备做出手掌朝正下方进行移动的机能性,因此可以判定它们是专性的二足步态。
另外,鹦鹉嘴龙的手掌也与其他鸟臀类的结构不同,它们的前肢只有四个指骨并长有利爪和长脚趾,显示出这是一群非常善于挖掘觅食的动物。但由于它们的前肢过短应该无法做出挖掘的动作,因此反而是依靠后肢进行挖掘,前肢仅仅起到送食和抓握的辅助功能。

有趣的是,鹦鹉嘴龙会通过吞食石子的方式来协助磨碎胃部等消化系统中的食物。目前发现的化石中,在腹部位置就有超过50颗胃石化石,推测跟大部分现代鸟类一样,这些胃石应该储藏在它们的砂囊中。
在它们的某些骨骼残骸中,还发现了许多坚硬的植物根茎和果实外壳,从而进一步佐证了鹦鹉嘴龙的食性和进食方式:不同于后期角龙类更加弯曲的喙部,它们的喙嘴显得更加平直而没有曲度,因此在进行咬合时只能采取后缩下颌再用嘴部后方切断植物后吞咽,并借助胃部的胃石辅助消化。
“毛发or羽毛?”
长久以来,羽毛似乎都是部分兽脚亚目恐龙(含鸟类)独有的衍征,在其他恐龙演化支上并未发现类似的结构。然而一个发现于中国辽宁省义县的鹦鹉嘴龙化石身上,却有着不同寻常的覆盖物,再次揭示出演化的神奇。
同大部分角龙科成员的化石一样,这个标本的皮肤压痕显示出其体表大部分也是由不规则排列的鳞片构成。然而耐人寻味的是,在背部至尾部末端处有一排明显的中空管状刺毛被完美的保存下来,长度最大接近16cm。
将其与同一地层出土的兽脚类原始羽毛对比可以发现,鹦鹉嘴龙尾部的管状鬃毛与兽脚亚目的原始羽毛并非是同源演化所致,两者应该是各自独立演化出现的。由于这些鬃毛仅分布在尾部的狭小区域因此它并不具备和羽毛一样的调节体温的作用,很可能只是一种视觉展示工具,用于求偶吸引伴侣。
而分属于兽脚亚目和鸟臀目不同种类的多个恐龙种群都各自平行独立演化出了羽毛、鬃毛等毛发结构,也说明了物种在演化上的复杂性。




想想看,在鹦鹉嘴龙生存的年代,四周强敌环伺,它们又缺乏后期角龙类的头盾和尖角,体型也不占优势,或许它们在捕食者来犯之时,会通过竖起、抖动这些管状鬃毛起到阻吓敌人的作用达到一定的防御性功用也不是不可能。
“纽带”
●【原角龙科 Protoceratopsidae 】:拉丁学名释义是“首个有角的脸”的意思,属于早期的冠饰角龙类。其下有一个模式种:安氏原角龙(Protoceratops andrewsi)
【生存年代】:上白垩纪坎潘阶;分布于蒙古国和中国,生存区域与伶盗龙重合。

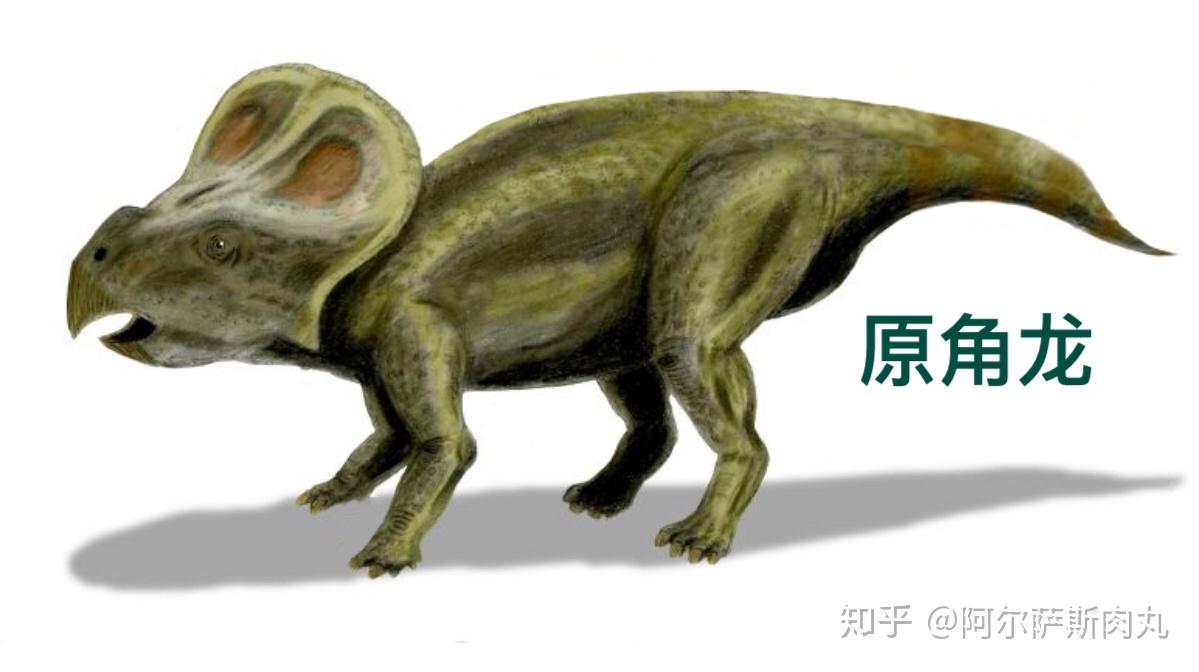
相比之前的鹦鹉嘴龙,原角龙各项生理特征都更加先进但依旧比后期角龙科原始,这体现在它们的四肢比例明显更善于快速奔跑且已转变为完全的四足步态、体型逐渐大型化其最大体长约有1.5-2米左右,大小接近于绵羊并演化出用于防御、保护颈部肌肉的头盾但依旧缺乏角。它们的头盾由颅顶骨和部分鳞骨特化而成,大小因个体差异略有不同。研究认为,它们头盾的差异性可能和两性异形有关。


根据大量发现的沉积化石标本推测,原角龙是典型的群居性动物,成年个体体重能达到180公斤。由于它们的生存区域与蒙古伶盗龙几乎完全重合,由此可见集体防御的策略能够更加有效的抵抗来自捕食者的入侵,毕竟它们的天敌伶盗龙也是采取集体狩猎的模式。
作为一种相对小型的角龙,原角龙的头颅占据了身体的大部分。而它们粗壮结实的嘴部肌肉,具备极佳的咬合力,非常适合咀嚼坚硬的植物根茎;与此同时它们的有着一个大型的眼眶,直径约有50mm,后方则是下颞孔。这表明原角龙有着出众的视力且夜视能力应该也不弱,而敏锐的视觉在帮助它们防范躲避捕食者攻击时能起到举足轻重的作用。
同鹦鹉嘴龙一样,目前也并不认为原角龙是后来更进阶的新角龙类的祖先。
●【祖尼角龙属 Zuniceratops】:学名意为“来自祖尼部落的长角面孔”。
【生存年代】:晚白垩纪中土伦阶;分布于美国新墨西哥州,极有可能是角龙科的祖先。




祖尼角龙的化石出土于1996年,由美国古生物学家Douglas G. Wolfe在新墨西哥州发现。这是一种身长有3-3.5米、肩高1米、体重达到100-150公斤的角龙类。它们是目前已知最早的有额角结构的北美洲角龙科成员,但其头盾化石显示依旧缺乏后期角龙的缘骨突(eroccipital)。
值得一提的是,祖尼角龙可以说是连接原始的早期角龙科成员(如:原角龙)和后期的大型进阶型角龙类(如:三角龙)之间的一个纽带。作为过渡演化物种的典型代表,祖尼角龙的发现进一步支持了角龙下目起源于北美的理论。
“源起何处?”
传统的观念一直认为角龙超科起源于北美洲。而在2009年,随着位于中亚的乌兹别克斯坦图兰角龙化石被发现(这是唯一一种生存于亚洲的角龙科恐龙),则预示着角龙科的起源地也许在亚洲大陆。
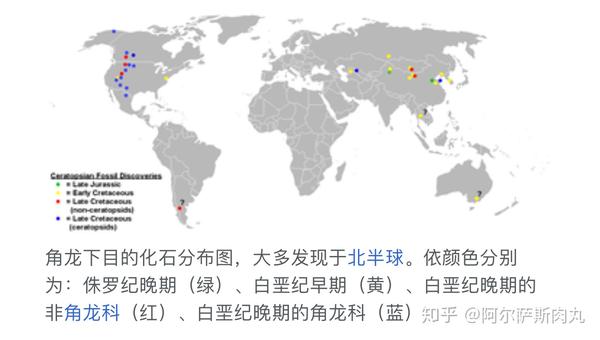

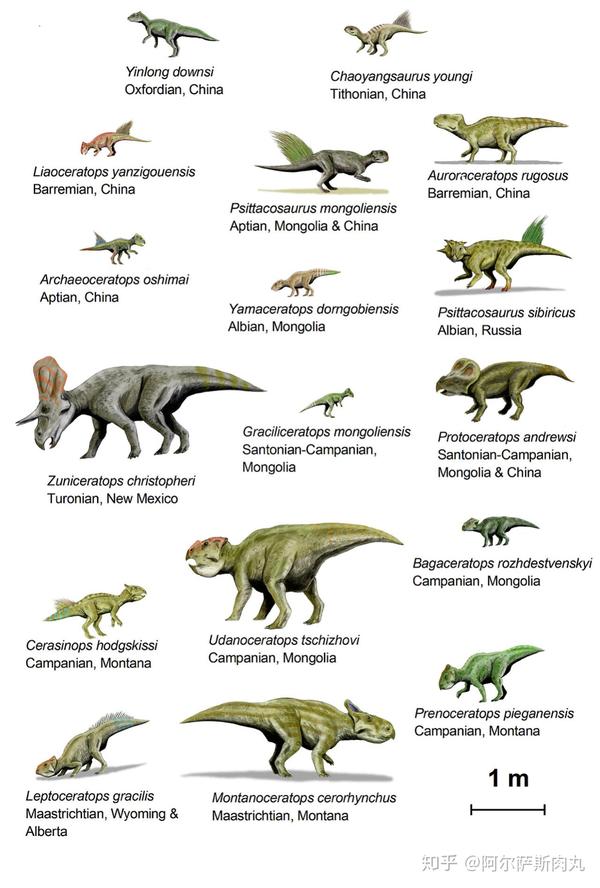
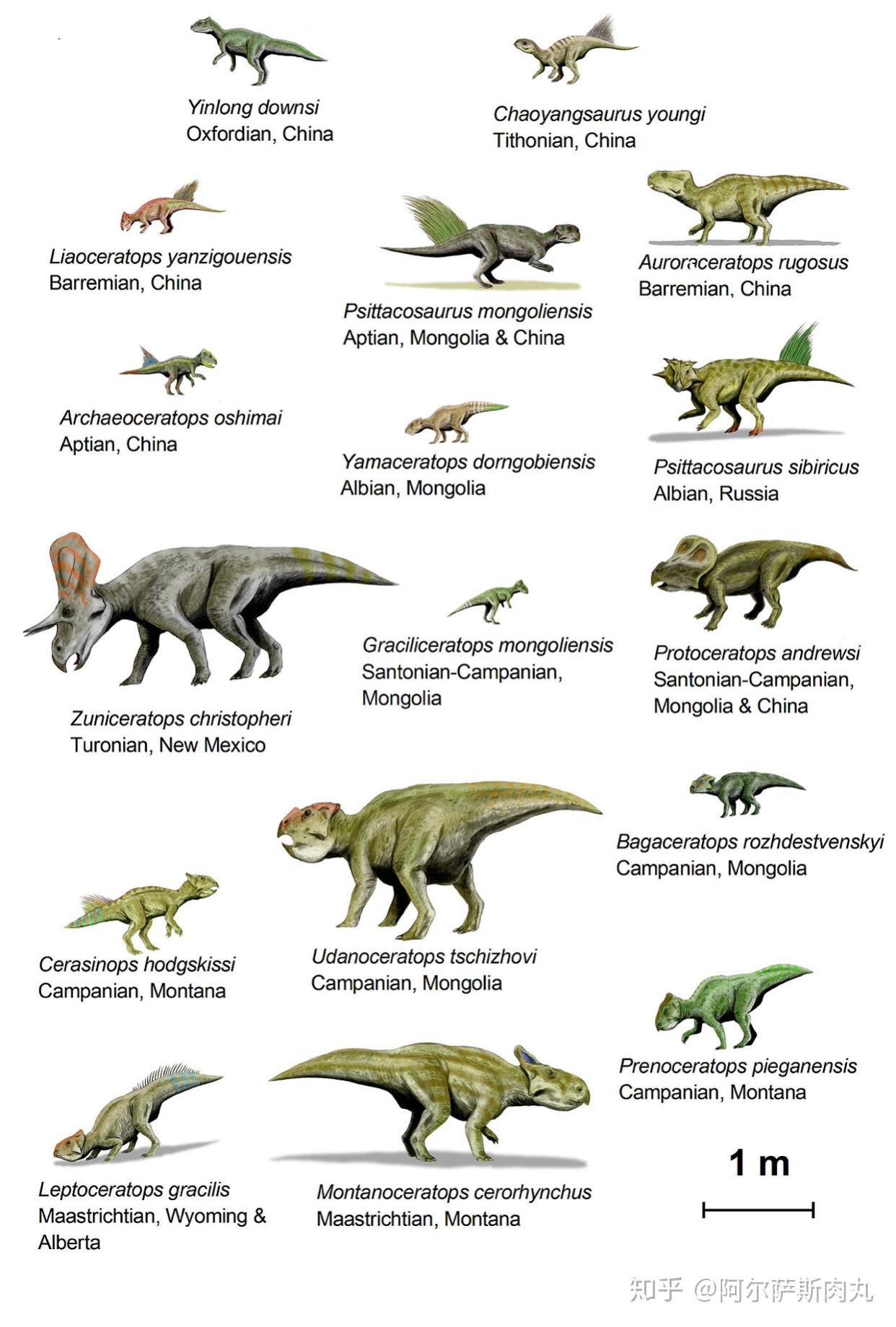
有趣的是,虽然早期的角龙下目成员化石均发现于亚洲,但在距今1亿1200万年前的北美阿尔比阶地层中也出土了不少新角龙类的化石。显示出在中白垩纪时期,一部分角龙下目横跨了现今的白令海峡到达北美洲并迅速开始扩散演化成为优势种群,但祖尼角龙似乎是北美西部地区的特有种。
不论是起源于亚洲还是北美,角龙下目都凭借出众的适应力迅速成为了当时陆地最成功的植食性动物种群。它们同新崛起的开花类植物一道,共同谱写出末日来临前的绚丽篇章。
“装备升级”
●【尖角龙亚科 Centrosaurinae】,学名释义“带尖刺的蜥蜴”。
【生存年代】:距今7650-7550万年前的上白垩纪,加拿大/美国。


跟早期小型、两足行走的鹦鹉嘴龙前辈不同,后期以尖角龙为代表的角龙随着体重的增加和重心的转移逐步适应演化为具有四足步态、面部额部有夸张角状结构和粗壮颈部头盾的庞然大物。
它们从侏罗纪晚期开始到白垩纪晚期的大型化之路上可谓一骑绝尘:体长从最初的1米演变到9米、体重更是从23公斤跃升到5.6吨;其头颅顶骨、方轭骨也更加向后弯延伸形成明显的头盾;颧骨往两侧更加突出使得从上方观察时呈现出明显地三角形结构。
而作为一种中型植食性恐龙,尖角龙体长一般可达6米,四肢粗壮。所有尖角龙族成员都有着狭长的脸部和突出的鼻角,在双目上方也有一对略微向上弯曲的小型额角;头盾较短粗附有孔洞,周围分布有细小的角突。


由于独特的头盾和角饰,角龙类成为恐龙大家族中最具辨识度的成员。而有关它们的角和头盾的具体功能,长久以来一直是学界争论的热点话题。
普遍理论认为:这是角龙类用于抵御捕食者、内部雄性争斗的工具。然而一项2009年的研究报告在比较了三角龙和尖角龙的颅骨损伤程度后发现,这些创伤都是同类打斗行为导致的。也就是说这些利器用于对抗肉食性掠食者的可能性较小。
而相对三角龙实心无孔洞的厚实头盾,尖角龙的头盾相对薄弱无法起到有效的防御作用,因此推测其头盾充当视觉辨识作用、吸引异性交配的可能性更大。
●【戟龙属 Styracosaurus】学名意为“带刺盾的蜥蜴”。
【生存年代】:距今7650-7500万年前的白垩纪坎潘阶,加拿大/美国。
再说说同样属于尖角龙亚科的戟龙,它可算得上是仅次于三角龙的热门物种。它们最与众不同的地方就是那往两侧和上方延伸出的四至六对尖角和夸张的头盾,除此之外戟龙的鼻角也非常修长,约有60公分长、15公分宽且向内弯曲。
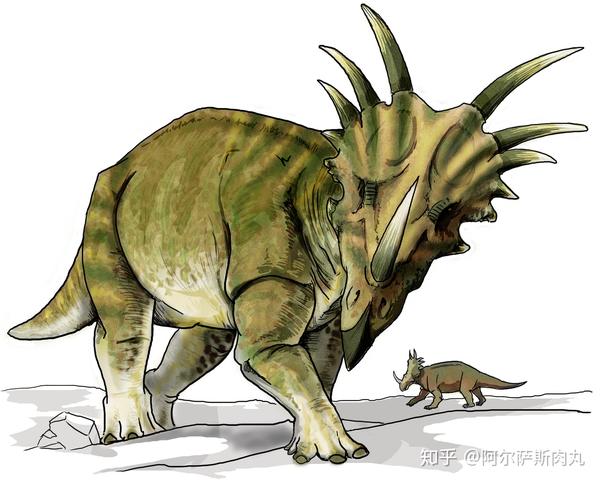



作为一种大型的角龙类,戟龙体长可达5.5-6米、身高1.8米、体重有2.7吨;有着短粗结实的四肢,脚趾上有被角质层覆盖的蹄状爪类似于现代的犀牛;较短的尾部和强壮高耸的肩部可用于打斗(如争夺领地和抢夺配偶权);硕大无比的头颅上有着弯曲的头盾和尖角,在头盾边缘还有许多尖刺一样的凸起;嘴部前端是无牙的角质喙便于咬碎坚硬的植物果实外壳和根茎……所有的这些装备都显示出戟龙是一种高度特化的进阶型角龙。




作为与尖角龙属关系最亲的近亲,戟龙也有不少独到之处:由于相对粗短的四肢,它们主要以低处的植被为食,但也能通过厚实的肩部和头角撞击较高的植物使其折断从而获取更充足、多样化的食物;另外在进食模式上,由于戟龙的喙嘴比较狭窄修长说明这更加适合拉扯、撕扯食物而非咬合。
需要说明的是,在角龙科的齿系中,上方较老的牙齿会被下部更新的陆续取代且这一过程终生不断。且相对同样具备卓越咀嚼能力的鸭嘴龙科而言,角龙类的牙齿更适合咬合,而鸭嘴龙科则更适用于磨碎。因此在食性选择上,角龙类主要以棕榈科植物或苏铁为主,通过撞到树干获取新鲜的叶片或挖掘多汁根部为食。
“巅峰”
●【三角龙属 Triceratops 】: 拉丁学名为“三角的蜥蜴”之意。属于开角龙亚科(Chasmosaurinae)成员。
【生存年代】:晚白垩纪马斯特里赫特阶;距今约6800-6500万年前的北美洲。
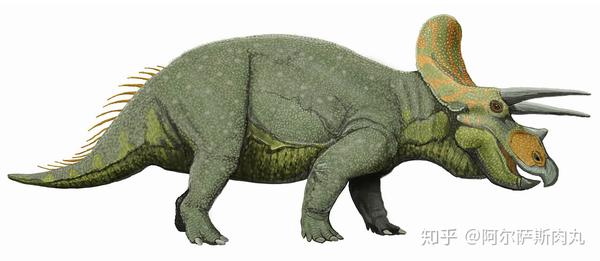
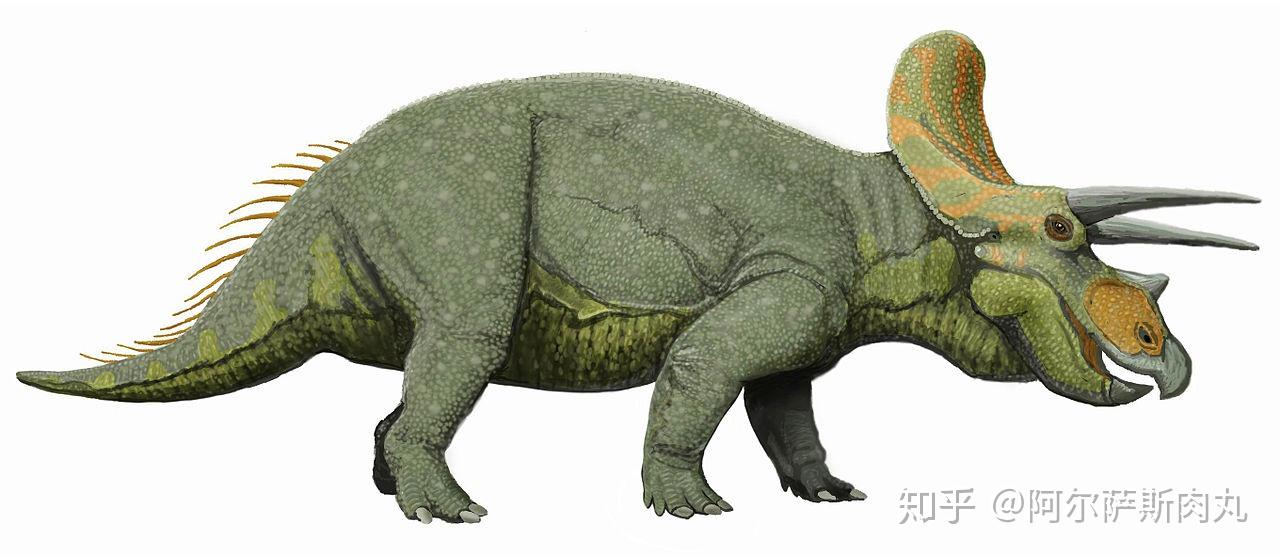
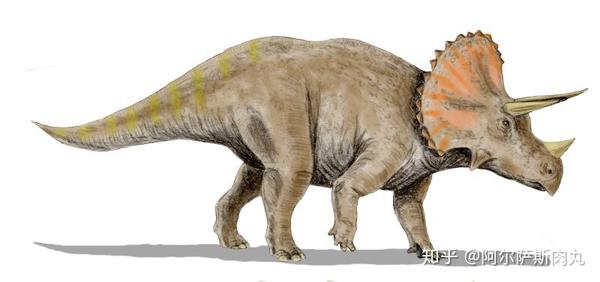

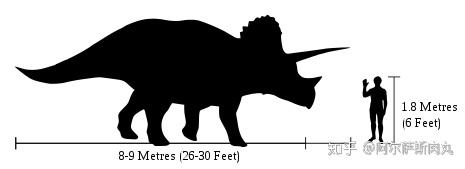

作为恐龙家族中最晚出现的恐龙之一,同时也是中生代白垩纪晚期种群数量最多的三角龙,其化石数量之多、分布之广泛更使其成为了白垩纪晚期的指标性物种化石。
它们凭借显眼的三根利角和壮硕无比的体型一跃成为大众流行文化中最受欢迎的恐龙。再加上三角龙的化石分布区域同暴龙科几乎完美重合,这也为研究了解两个物种间的竞争演变和古地理学生态提供了绝佳的参考样本。
从外形上看,三角龙拥有着所有陆地动物中最大之一的头颅,其头盾最长可超过2.5米,占整个身体比例的1/3。相比其他具有角盾的大型角龙而言(如:尖角龙),三角龙的骨质头盾则是完全实心的无孔洞结构。以上种种性状都显示出三角龙代表了角龙科恐龙演化的巅峰。




再对比三角龙的四肢可以发现,与鸟臀目-装甲亚目和蜥臀目-蜥脚亚目的大型植食性恐龙不同,三角龙的前后肢脚趾依旧保留了许多原始的特征:当它们行走时,前脚掌的脚趾是朝前并往两侧撑开而非像蜥脚亚目那般脚趾向前掌心向后,这样的步态在兽脚亚目和早期两足行走的原始鸟臀目身上都能见到。
另外,与现代的大象和犀牛不同,三角龙前脚掌的五个趾头只有三趾起到支撑体重的作用,第三和第四趾已退化且并未形成蹄状肢。






与所有的角龙一样,三角龙的牙齿也是终生生长并不断替换的:它们通常具有432~800颗牙齿,这些牙齿排列成齿列,每列由36-40个齿群组成;上下颌两侧各有3-5列齿群。这些牙齿采取垂直的方向切割食物,根据其磨损程度推测出,它们的食谱上主要是苏铁和棕榈这样的大体积粗纤维植物,偶尔也会啃食蕨类植物。
防御迷思


在很多影视作品或复原资料中,我们时常能够看到三角龙用尖角对进犯的暴龙予以猛烈还击的场面,这一幕几乎也成为了角龙科与暴龙科对决的经典。然而,三角龙的角和头盾真的足以抵御暴龙这样的顶级掠食者的袭击吗?
有关三角龙的头饰功能目前的假说理论主要有两种:1.用于战斗、抵抗外来攻击;2.求偶过程中的视觉展示。
而目前,越来越的证据更加倾向于后者。

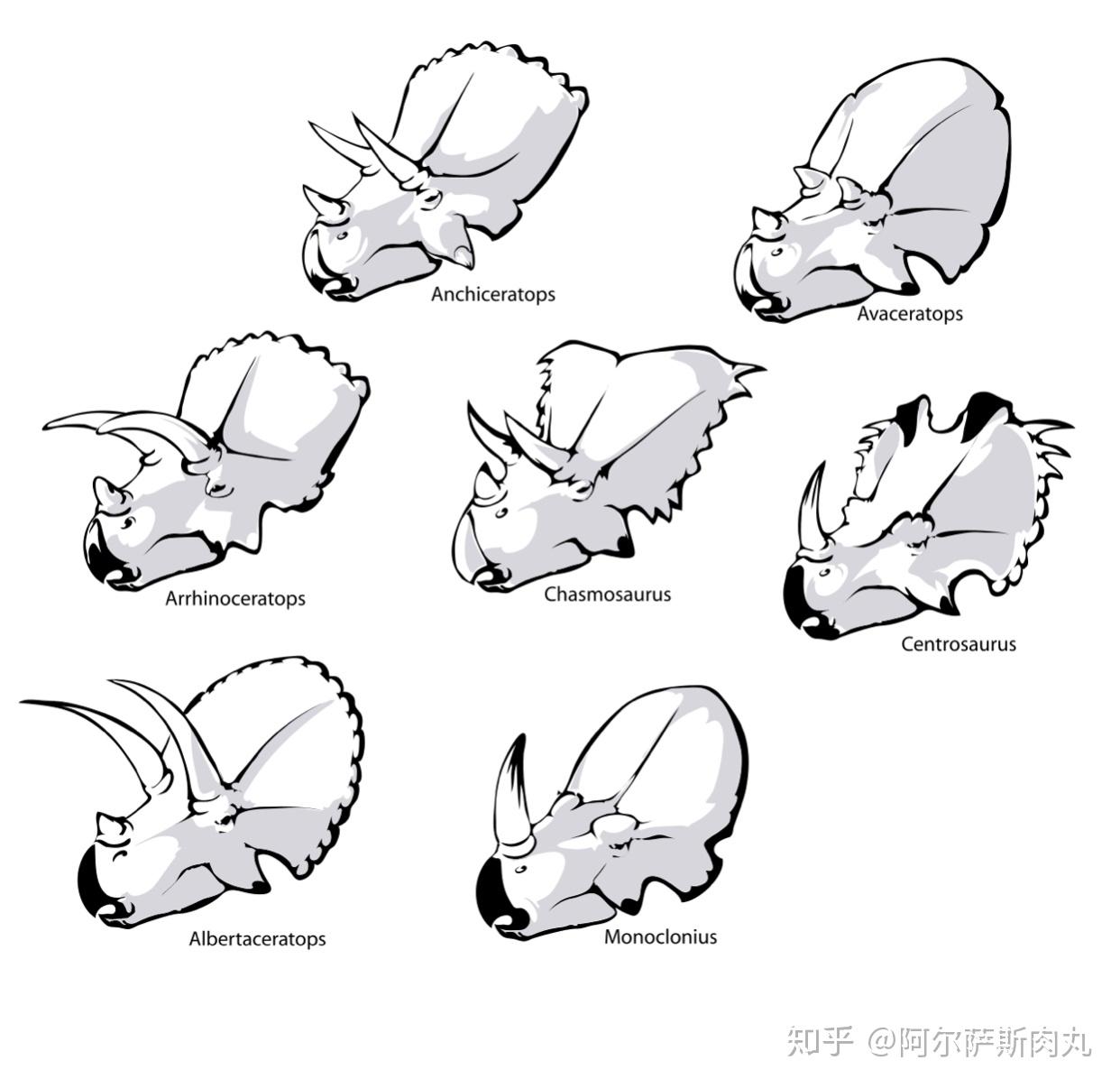




在早期的研究中一直认为,三角龙会使用它锋利的角和坚实地头盾同暴龙这样的掠食者进行战斗,这个理论的出现最早可以追溯到1917年,由查尔斯.斯滕伯格首次提出。
他根据当时的化石证据指出,暴龙会以三角龙为食,并在一个断裂的三角龙额角和鳞骨上发现了暴龙牙齿的咬痕及痊愈后的迹象,表明这样的攻击事件是在三角龙存活时发生的而非食腐行为所致。而后的一些古生物学家如彼得.道森则更进一步假设出三角龙会将前肢抬高后用头部的角对准暴龙腹部进行刺杀,这也成为后续很多复原资料中采用的依据来源。


然而,事实真相果真如此吗?
2005年,英国BBC的《The truth about Killer Dinosaurs》(恐龙真凶/恐龙杀手的真相)节目组进行了三角龙如何抵御暴龙袭击的模拟测试,或许能为我们揭开其中的一些奥秘。
节目组制作了一个同等比例复原的三角龙头颅模型,并以24KM/h的速度快速冲撞向人工模拟的暴龙皮肤,结果显示:三角龙的额角能够刺入皮肤但无法深入刺穿到肌肉,且颅骨的前端出现裂痕。这意味着,三角龙其实不大可能冒着头颅和尖角断裂的风险用高速冲撞的策略进行防卫。而更可能采取坚守策略近距离的用角和头盾对来犯的天敌进行反击。
也就是说,大部分电影中三角龙横冲直撞如犀牛一般冲向掠食者的场景,在现实中是不可能上演的。


那么,除了用于防御之外三角龙的头饰还有哪些其他的功用呢?
在比较了三角龙/尖角龙的颅骨损伤比率后可以发现:三角龙更加频繁的使用头角进行打斗;而尖角龙的头盾上则很少发现类似的伤痕说明它们更倾向于用头饰进行展示而非打斗。
而戟龙那特化的大型头盾很可能起到有效增大身体表面积达到散热调节体温的目的,功能类比大象的耳朵。




最近的研究显示,三角龙尤其是雄性之间为了争夺领地统治权和交配权以及吸引配偶,会用头角进行碰撞,就如同今天非洲草原上的角马一样。
在一些三角龙的颅骨化石上都发现了由冲撞形成的孔洞、创口痕迹,表明三角龙同类之间互相争斗、打斗而留下伤痕推论是合理存在的。而像戟龙这样的夸张头盾甚至有可能会通过毛细血管充血的方式使其呈现出不同的颜色和图案,达到吸引异性获得优先交配权的目的甚至恐吓捕食者的目的。






自1961年Davitashvili首次提出“头饰/头盾用于展示”的理论至今,这一学说目前也越来越得到学界的赞同支持。由此可见,通过对比现代具有同样衍征的动物将角状物作为视觉辨识物使用的案例,我们可以合理佐证推测出:在不同种类的角龙种群上,借助形态各异的装饰性头饰来增强其在求偶等社会性行为中的优势地位现象是客观存在并合理的。
【结语】


从帝龙到暴龙、从鹦鹉嘴龙到三角龙、恐龙阵营中的两大种群展开了一场从侏罗纪绵延至白垩纪的漫长且势均力敌的演化逆袭战,你攻我守打得火热。然而这场军备竞赛最终还是被一颗落在尤卡坦半岛的陨石彻底打断,提早落幕划上了句号。
对于我们人类而言,回顾它们令人唏嘘的演化史时,就更加明白了生命的渺小和演化的不确定性。
地球在一次次周而复始的大灭绝-大复苏之间不断切换,没有谁是永恒的王者。
没准,下一颗可能毁灭掉人类文明的陨石就正在赶来的路上呢……
【THE END】
→下期预告>>>“白垩传3[沧海浮沉]:沧龙兴衰录”


◎【词条释义】:新角龙类/冠饰角龙类
定义:所有比鹦鹉嘴龙科更衍化进步的角龙下目。其中冠饰角龙类首先发展出头盾结构,其颈部前端的几个脊椎骨用于固定和支撑增重后的颅骨是它们最显著的特征。通常有三个演化支被系统承认,分别是:
1. 原角龙科:包括原角龙及近亲成员,全部分布于亚洲;不过它们并非是后来一系列角龙科的直系祖先。
2. 纤角龙科:包括纤角龙与其相关的亲缘演化支,主要分布于北美洲;
3. 角龙超科:包括祖尼角龙和近亲角龙科。其中囊括了以三角龙为代表的所有大型北美洲角龙下目种群,如尖角龙亚科和角龙亚科。
*注:部分图片及文字资料引述自维基百科中文版/英文版/ DeviantArt,原作者已打水印或署名的图片不再单独备注。
